Autores:
Katherine Calderón1, Aldo Rojas-Neyra1, Ángela Montalvan-Avalos1, Gisela Isasi-Rivas1, Brigith Carbajal-Lévano1, Gloria Guerrero-Fonseca1, Julio Ticona1, Luis Luján-Valenzuela1, Suly Montoya-Ortiz1, José Goggin-Ortiz1, Manuel Criollo-Orozco1,Edison Huaccachi-Gonzáles1, Luis Tataje-Lavanda1, Doris Villanueva-Pérez1, Milagros Lulo-Vargas1, Astrid Poma-Acevedo1, Freddy Ygnacio1, Ricardo Montesinos-Millán1, Manolo Fernández-Sánchez1, Manolo Fernández-Díaz1
1Laboratorios de Investigación y Desarrollo (I+D), Farmacológicos Veterinarios S.A.C (FARVET SAC), Chincha alta, Ica, Perú. 24/01/2023
INTRODUCCIÓN
La influenza aviar (AI, por sus siglas en inglés: Avian Influenza), es una enfermedad causada por el virus de la influenza aviar (AIV, por sus siglas en inglés Avian Influenza Virus), es de gran importancia para la industria avícola, debido a las enormes pérdidas económicas, eliminación de millones de aves de crianza comercial, restricciones al comercio internacional, y además de tener un alto riesgo zoonótico poniendo en peligro la salud humana (1).
El virus de la AI es miembro del género Orthomyxovirus, de la familia Orthomyxoviridae según el Comité Internacional de Taxonomía de Virus (ICTV, por sus siglas en inglés: International Committee on Taxonomy of Viruses). El virus tiene un genoma de ácido ribonucleico (ARN) segmentado, de cadena simple de sentido negativo y contiene 8 segmentos que codifican: polimerasa básica 1 (PB1), polimerasa básica 2 (PB2), polimerasa ácida (PA), hemaglutinina (HA), nucleoproteína (NP), neuraminidasa (NA), matriz 1 (M1), matriz 2 (M2). IAV también expresa muchas proteínas no estructurales (NSP), como NS1, que no están empaquetadas en la partícula viral de AIV, sino que solo se expresan en las células huésped infectadas. Recientemente, se han descubierto muchos NSP, incluidos PA-X, PB1, PB1-F2, PB1-N40, PA-N155, PA-N182, M42 y NS3, todas estas proteínas juegan un rol crucial en la patogenicidad del virus. Las proteínas hemaglutinina (HA, por sus siglas en inglés: haemagglutinin) y la neuraminidasa (NA, por sus siglas en inglés: neuraminidase) son glicoproteínas de superficie importantes para la infectividad del virus. La proteína HA es responsable de la unión del virus hacia la célula huésped y juega un rol importante en la respuesta inmune humoral. La proteína HA es sintetizada como una proteína precursora (HA0) en células infectadas y requiere proteasa de las células hospedadoras para clivarse en forma activa (HA1 y H2).
La proteína NA, influye en la liberación y diseminación de viriones por remoción de ácido siálico que se une hacia la proteína HA, los anticuerpos contra la NA inhiben la unión o fusión entre el ácido siálico ubicado en el receptor celular e inhibe la fusión entre el virus y la membrana celular de la célula hospedadora. Asimismo, las propiedades antigénicas del virus AI esta íntimamente relacionado con sus glicoproteínas de superficie como la HA y la NA, se han identificado 16 subtipos de HA (H1-H16) y 9 subtipos de NA (N1-N9) aislados de aves acuáticas (2).
Diversas especies de aves han demostrado susceptibilidad ante la infección del virus AI; aves acuáticas, aves comerciales, pavos etc. La alta virulencia del virus de la influenza aviar H5N1 produce una enfermedad aguda con alta mortalidad en ponedoras, pavos, y otras aves de interés económico.
El virus de AI por ocasionar una alta mortalidad en aves domésticas y silvestres está siendo considerada una prioridad emergente por la OIE y de importancia por la Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO, por sus siglas en inglés: Food and Agriculture Organization), y la organización Mundial de la salud (WHO, por sus siglas en inglés: Wordl Health Organization) por ser una enfermedad zoonótica.
La situación actual del virus de la AI de alta patogenicidad, especialmente del subtipo H5N1 linaje asiático, ha cambiado la ecología y epidemiología de la enfermedad, requiriendo la generación inmediata de nuevos conocimientos en temas relacionados a su patogénesis y dinámica de transmisión.
En Perú, solo se había registrado hasta hace unos meses atrás subtipos de AIV de baja patogenicidad (LPAI) por aislamiento viral a partir de muestras fecales de aves silvestres colectadas en zonas de la costa, que usualmente coincidía con los meses de verano donde se alcanzaba una alta concentración de aves migratorias silvestres (3–6). Perú era hasta hace poco un país libre de AI, sin embargo, con el ingreso de aves migratorias en noviembre del 2022 el SENASA-PERU ha reportado la presencia del virus H5N1 en pelicanos, aves de crianza casera y comerciales (7) (Resolución Jefatural Nº 0173-2022-MIDAGRI-SENASA 23 de noviembre de 2022).
Hoy en día para el control del virus de AI, existen vacunas inactivadas o las llamadas vacunas convencionales que son preparadas a partir de fluido alantoideo de huevos SPF inoculados y luego inactivado con beta-propiolactona o formol, que confieren inmunidad después de su administración y, para una protección efectiva en aves de larga vida, requieren múltiples dosis de refuerzo. Su aplicación y efectividad se ve limitada debido a que por lo general es importante el empleo de vacunas con cepas homologas (vacunas preparadas a partir de virus aislado en una epizootia), demostrando ser más eficaces en la producción temprana de anticuerpos y con altos títulos que correlacionan con una protección completa y efectiva en las aves vacunadas.
Otras vacunas como las vectorizadas para AI, se ha basado en la inserción del gen codificante HA en el genoma de un vector viral recombinante, el cual presenta las siguientes ventajas: I) induce inmunidad mucosal, humoral, y celular, II) puede ser administrado en pollos jóvenes e inducir una protección temprana, por ejemplo: Fowl poxvirus (vacuna fowlpox-AI-H5), Newcastle (rNDV) que expresa un gen H5 HA (rNDV-H5) y el HVT o herpes virus del pavo (rHVT-AIV H5) y II) puede ser diferenciado entre aves vacunadas e infectadas (1). Sin embargo, el uso de estas vacunas es limitada debido a que solo son eficaces en ausencia de anticuerpos contra AIV. La vacuna rHVT-AIV H5 superó la interferencia de anticuerpos maternales contra AIV y brindó protección contra HPAIV (8), Por otro lado, la vacuna fowlpox-AI-H5, no ha logrado provocar una respuesta inmunológica detectable. La elección del vector es muy importante para conseguir una buena protección (8).
Para superar estas limitaciones, diversas estrategias para expresar proteínas inmunogénicas de las proteínas de AIV en vectores vivos han sido exploradas (9). El más común vector vacunal utilizado es el Meleagrid herpesvirus 1 (MeHV-1) o llamado Herpesvirus de pavo (HVT). El HVT pertenece al serotipo 3 del virus de la enfermedad de Marek (MDV). Es un alfaherpesvirus no patogénico en pollos, que contiene un genoma de ácido desoxirribonucleico (ADN) de doble cadena. El HVT es ampliamente utilizado como vector vacunal contra MDV y es considerado un candidato vacunal estable, debido a su habilidad de persistir en la infección, e incluso en la presencia de anticuerpos maternales, además decrece la diseminación viral e induce inmunidad humoral y celular de larga duración (siendo detectado por más de 50 semanas de edad) con una sola dosis de vacunación que puede ser administrada al primer día de edad por vía subcutánea o a los 18 días de edad embrionaria (por vía in ovo) (10).
El HVT, ha sido ampliamente utilizado como vector viral para la expresión de antígenos virales de diversas enfermedades aviares tales como la enfermedad de Gumboro (IBD) (11), laringotraqueitis infecciosa (ILTV), chlamydia psittacia, la enfermedad de Newcastle (ND) (12) e influenza aviar (AI) (13, 14).
Recientemente la nueva plataforma de edición génica de Repeticiones Palindrómicas Cortas Agrupadas y Regularmente Inter espaciadas (CRISPR, por sus siglas en inglés: Clustered Regularly Interspaced Short Palindromic Repeat) más Cas9 (endonucleasa Cas9 de Streptococcus pyogenes) ha sido empleada en el desarrollo de nuevas vacunas vectorizadas en base al vector viral HVT, debido a su gran versatilidad de ser fácil, rápida, y económica.
En este estudio, nosotros demostramos la eficacia protectora de la vacuna viva recombinante (rHVT-AIV H5) y la vacuna inactivada de AIV subtipo H5N1 (vacuna autógena) aplicadas en tres diferentes programas de vacunación como una estrategia preventiva o de emergencia para el control del virus de AI altamente patógena H5N1.
MATERIALES Y MÉTODOS
- Animales
Aves libres de patógenos específicos (SPF, por sus siglas en inglés), Charles River Avian Vaccine Services, Norwich, USA, white leghorn de 1 día de edad.
- Células y virus
Se utilizaron células de fibroblasto de embrión de pollo (CEF, por sus siglas en inglés: Chick embryo fibroblasts) extraídas de 10 días de edad de huevos embrionados SPF. También, se emplearon células de riñón canino (MDCK, por sus siglas en inglés: Madin-Darby canine kidney). Todas las células fueron mantenidas en DMEM/F12 (DMEM, por sus siglas en inglés: Dulbecco’s Modified Eagles medium) (Thermo Fisher Scientific) suplementado con 5% de suero fetal bovino inactivado (FBS, por sus siglas en inglés: fetal bovine serum) (Hyclone) y 1X de antibiótico-antimicótico (Thermo Fisher Scientific), a 37 °C, bajo 5% CO2 de atmósfera.
El wild-type HVT (wtHVT) cepa FC 126, fue utilizado para la generación del recombinante rHVT-AIV H5 en células CEFs por la tecnología CRISPR/Cas9 y NHEJ (nonhomologous end-joining).
La cepa viral de AIV H5N1, fue previamente identificada y secuenciada por FARVET, cuyo genotipo correspondió a H5N1 clado 2.3.4.4b, denominado cepa VFAR-140. Esta fue utilizada para la preparación de la vacuna inactivada de AIV H5N1, y asimismo utilizada como cepa de desafío de AIV H5N1 para demostrar la eficacia de las vacunas de este experimento.
- Obtención de la vacuna rHVT-AIV H5
Generación del recombinante rHVT-AIV H5 que contiene el gen HA de AIV H5
El virus recombinante rHVT-AIV H5, fue obtenido por la tecnología CRISPR/Cas9-NHEJ utilizando los plásmidos y sgRNAs: pGEM-sgA-GFP-AIV, px459v2.0-sgRNA-sgA, y px459v2.0-sgRNA1-HVTUL55-56.
El marco de lectura abierto (ORF, por sus siglas del inglés: Open Reading Frame) de la proteína HA del virus de influenza aviar HPAIV A (A/Broiler chicken/Giza/VRLCU/2019) (H5N8), subtipo H5 de la cepa H5N8 (Número de acceso al GenBank: MT256068.1) corresponde a una cepa de alta patogenicidad, el sitio de clivaje del gen HA5, fue modificado a una secuencia de sitio de clivaje de baja patogenicidad, eliminando 3 residuos: lisina-arginina-arginina “KRR” (posición 340-343) y reemplazando un residuo de lisina (K) por una treonina (T) (posición 344). Esta secuencia fue sintetizada por GenScript y subsecuentemente clonada y finalmente insertada en la región intergénica no codificante UL55-56 del genoma HVT por la tecnología CRISPR/Cas9-NHEJ (Figura 1).
La correcta inserción y orientación del casete de expresión HA de AIV H5, fue verificado por PCR con cebadores específicos (Tabla 1)
Figura 1. Flujograma de trabajo para la construcción y recuperación del virus recombinante rHVT-AIV H5 por CRISPR/Cas9-NHEJ.
Tabla 1. Lista de cebadores y de sgRNAs utilizados en este estudio para la generación del rHVT-AIV H5

Construcción y recuperación del virus rHVT-AIV H5
Los sgRNAs (sgRNA-sgA and sgRNA-UL55-56) fueron diseñados, sintetizados, y clonados en el plásmido px459v.20, el cual contiene el gen de la Cas9 de S. pyogenes. Los plásmidos resultantes fueron denominados px459v2.0-sgRNA-sgA y px459v2.0-sgRNA-UL55-56.
El plásmido donador pGEM-sgA-GFP-AIV contiene el casete de expresión del GFP reportero el cual se encuentra flanqueado por dos secuencias de LoxP para su separación del casete AIV de expresión. Los dos casetes fueron flanqueados por sitios de sg-A para introducir un corte deseado o clivaje, para la liberación del casete e integración en la región intergénica UL55-56 del genoma wtHVT. La endonucleasa Cas9 promueve la inserción del casete GFP y AIV en la región intergénica UL55-56 del genoma del wtHVT (Figura 2).
Para la generación del virus rHVT-GFP-AIV, las células CEF fueron co-transfectadas con los plásmidos pGEM-sgA-GFP-AIV, px459v2.0-sgRNA-sgA, y px459v2.0-sgRNA-UL55-56. A las 24 horas post-transfección, las células fueron infectadas con el virus wtHVT con una multiplicidad de infección (MOI, por sus siglas del inglés: multiplicity of infección) de 0.01. Las placas que expresan GFP fueron visibles a los 3 días post-transfección/infección, demostrando con éxito la inserción del casete. El virus recombinante fue aislado en tres rondas de purificación por aislamiento de placas de lisis y por Cell Sorter.
Posteriormente, la remoción del casete reportero GFP, fue realizado con el tratamiento de Cre-recombinasa (pcDNA3.1-Cre) a través de sistema Cre-LoxP, las células infectadas con el virus rHVT-GFP-AIV fueron co-transfectadas nuevamente con el Cre-recombinasa, la aparición de las placas de lisis sin emisión de fluorescencia fue visible a los 4 días post-transfección/infección (Figura 1). La completa remoción fue confirmada por PCR convencional utilizando cebadores específicos HVTUL55-2F y HVTUL56-2R (Tabla 1). Finalmente, el nuevo recombinante HVT fue denominado rHVT-AIV H5.
Estabilidad del gen HA en el genoma del virus recombinante rHVT-AIV H5
Las células CEFs fueron infectadas con el virus rHVT-AI VH5 durante 20 pasajes para determinar la estabilidad genética del virus mediante IFI, PCR y Western blot. El ADN viral fue analizado cada 5 pasajes por PCR donde detectamos la presencia del gen HA (Figura 3). Del mismo modo, cada 5 pasajes confirmamos la expresión de la proteína HA mediante Western blot (Figura 3B) donde detectamos una banda de ∼70kDa que representó la proteína precursora de HA (HA0), una banda de ∼50kDa que representó la subunidad clivada HA1, y una banda de ∼25kDa que representó la subunidad clivada HA2. El IFI demostró la expresión de la proteína HA en células infectadas con el virus recombinante en el pasaje 20 en CEFs (Figura 3C). Estos resultados demostraron que la presencia y la expresión del gen de la proteína HA del AIV no afectó la replicación del virus recombinante, confirmando la estabilidad del gen HA en el genoma viral del recombinante.
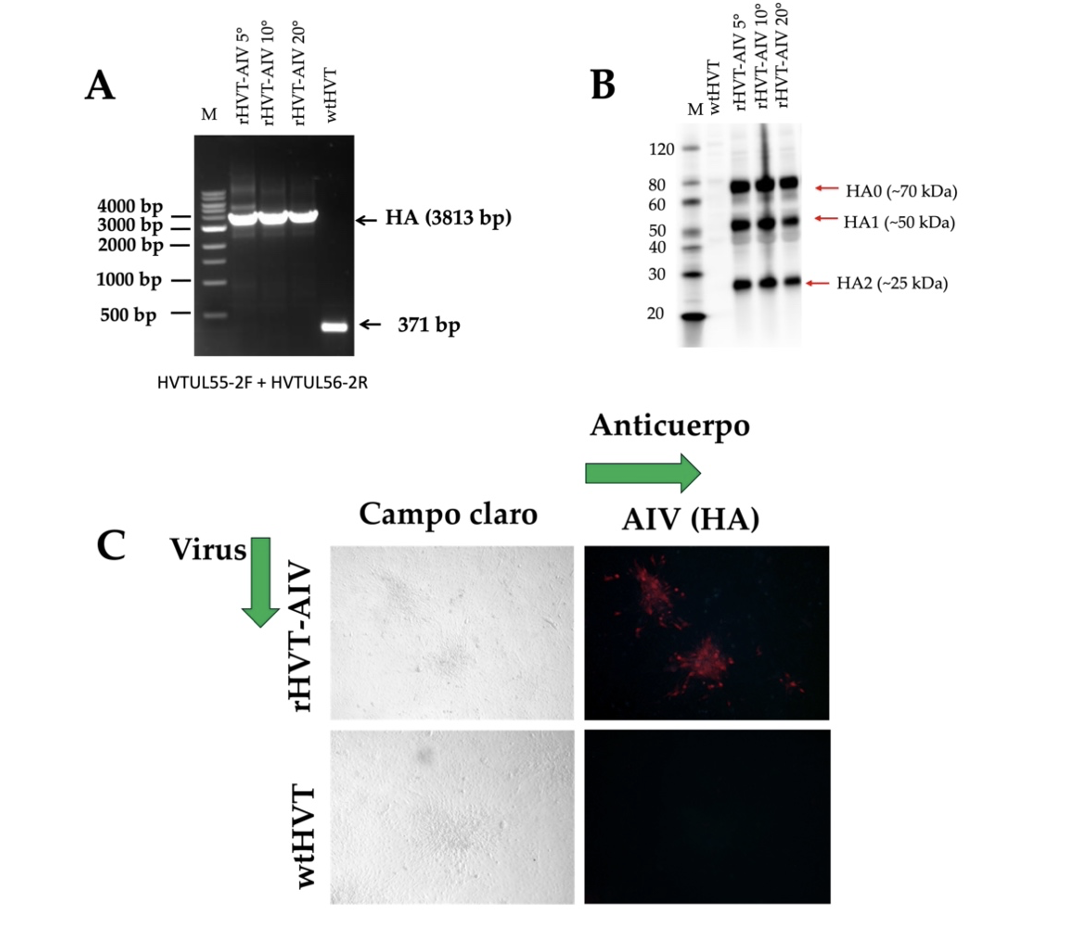
Figura 3. Muestra la evaluación In vitro de la estabilidad del gen HA en el genoma del virus rHVT-AIV H5. Las células CEF fueron infectadas con el virus rHVT-AIV H5 durante 20 pasajes. (A) Cada 5 pasajes la presencia del gen HA en el genoma del rHVT-AIV H5 fue evaluado mediante PCR convencional, el wtHVT fue utilizado como control negativo. la flecha negra indica el tamaño de la banda esperada. (B) La expresión de la proteína HA fue evaluado en células CEF infectadas con rHVT-AIV H5 mediante Western blot cada 5 pasajes, donde detectamos una banda de ∼70kDa que representó la proteína precursora de HA (HA0), una banda de ∼50kDa que representó la subunidad clivada HA1, y una banda de ∼25kDa que representó la subunidad clivada HA2. Como control negativo utilizamos células infectadas con wtHVT. La flecha roja indica el tamaño de la banda esperada. (C) La expresión de la proteína HA fue evaluado por ensayo de inmunofluorescencia indirecta (C) en células CEF infectadas con el rHVT-AIV H5, como control negativo se utilizaron células CEF infectadas con el wtHVT.
- Ensayo de inmunofluorescencia indirecta (IFI)
Células CEF se multiplicaron en placas de 12 pocillos y fueron infectadas con una MOI de 0.005 con el wtHVT y rHVT-AIV H5. Después de 48 horas post infección (hpi), las células fueron lavadas tres veces con DPBS 1X y luego se fijaron con 4% de para formaldehído por 15 minutos (min) a temperatura ambiente. Las células fueron incubadas con el anticuerpo primario en una solución de 5% albumina de suero bovino (BSA, por sus siglas en inglés: Bovine serum albumin) en DPBS por 1 h a temperatura ambiente. Un anticuerpo de pollo anti MDV (Charles River Avian Vaccine Services, Norwich, USA) fue utilizado como control positivo para la detección de la infección de HVT, y el anticuerpo de conejo contra la proteína HA de AIV H5N8 (Sino Biological) fue utilizado para la detección de la expresión de la proteína HA. Después de los tres lavados con DPBS, las células fueron incubadas con un anticuerpo IgG de burro anti-conejo Alexa Fluor 594 (Abcam, Cambridge, MA, USA) y un anticuerpo de cabra anti-IgY de pollo Alexa Fluor 405 (Abcam, Cambridge, MA, USA) respectivamente en DPBS con 5% BSA por 45 min a temperatura ambiente. Los resultados fueron observados utilizando un microscopio de fluorescencia ObserverA1 fluorescence microscope (Carl Zeiss, Germany). Las imágenes digitales fueron tomadas a 50X de amplificación y procesadas con la cámara de AxioCam MRc5 (Carl Zeiss, Germany).
- Análisis de Western blot
Para evaluar la expresión de la proteína HA de AIV. Células CEF se infectaron con el virus recombinante con una MOI de 0.01. A las 72 hpi, las células CEF fueron lisadas y analizadas por Western blot. El análisis de Western blot fue realizado utilizando un anticuerpo específico policlonal de conejo contra la proteína HA de AIV (Sino Biological) como anticuerpo primario. Por otro lado, se usó un anticuerpo IgG anti-conejo conjugado a HRP (GenScript Laboratories, Piscataway, NJ, USA) como anticuerpo secundario. Las visualizaciones de la expresión de las proteínas fueron detectadas utilizando una cámara de AZURE CCD (Azure Biosystems, Inc, Dublin, USA).
- Estabilidad genética in vitro de rHVT-AIV H5
La estabilidad genética fue evaluada en CEFs hasta el pasaje 20. El ADN viral fue extraído a partir de células CEF infectadas, cada 5 pasajes se verificó la presencia del casete e inserto HA, el cual fue confirmado por PCR, usando cebadores específicos (Tabla 1). La expresión de la proteína HA fue evaluada cada 5 pasajes por ensayo de Western blot, descrito previamente.
- Preparación de la vacuna inactivada/emulsificada de AIV subtipo H5N1.
La cepa viral de AIV H5N1, fue previamente identificada y secuenciada por FARVET S.A.C, cuyo genotipo correspondió a H5N1 clado 2.3.4.4b, denominado cepa VFAR-140.
Células MDCK, fueron infectadas con la cepa VFAR-140 de AIV. Luego de 36 hpi, el sobrenadante fue cosechado y titulado por ensayo HA dado como título 1:1024. Posteriormente, se inactivó el virus de AIV cepa VFAR-140 a una concentración de 0.1%, con formaldehído, se dejó incubando a 37°C por 48 h en agitación. Se comprobó la completa efectividad del proceso de inactivación, a través de 3 pasajes sucesivos en huevos embrionados SPF de 10 días de edad (Charles River Avian Vaccine Services, Norwich, USA). Asimismo, cada cosecha de un pasaje fue evaluado por el ensayo de HA, donde los 3 pasajes no mostraron títulos positivos.
Luego de la inactivación, se determinó la cantidad de antígeno HA en la vacuna inactivada de AIV H5N1, a través de un análisis de proteínas totales por SDS-PAGE. El perfil proteico nos indicó la presencia de diferentes proteínas estructurales de AIV, entre ellas la presencia del antígeno HA1 visualizado en la cuarta banda, con un porcentaje del carril total del 30.92 % correspondiendo aproximadamente a 185.52 µg de antígeno HA por ml (Figura 4). Luego se ajustó la concentración proteica para que cada dosis de la vacuna contenga una concentración mayor a 3 mg de HA. Finalmente, la vacuna fue emulsificada con el adyuvante montanide; este a su vez se homogenizó por alta presión con el disruptor celular (Emulsiflex).
- Diseño de estudio: Inmunogenicidad de las vacunas.
Un total de 80 aves SPF (obtenidas por incubacion de huevos SPF) de 1 día de edad fueron utilizadas en este estudio. Después del nacimiento todas las aves fueron trasladadas a un laboratorio BSL-3 en las instalaciones de FARVET S.A.C. Las aves para este estudio fueron divididas de forma aleatoria en cuatro grupos (Tabla 2, Figura 5) de 20 aves cada uno: Grupo I (n = 20) fueron inmunizados a 1 día de edad con la vacuna rHVT-AIV H5 con una dosis conteniendo 2500 unidades formadoras de placa (UFP)/ave en un volumen de 0,2 ml por vía subcutánea; el grupo II (n = 20) fueron inmunizados a 1 día de edad con la vacuna inactivada AIV subtipo H5N1 (Clado 2.3.4.4b) con una dosis de 3 mg de HA en un volumen de 0,2 ml por vía subcutánea; el grupo III (n = 20) fueron inmunizados a 1 día de edad con rHVT-AIV H5 conteniendo 2500 UFP y una dosis de la vacuna inactivada AIV subtipo H5N1(Clado 2.3.4.4b) con una dosis de 3 mg de HA en un volumen de 0,2ml; el grupo IV (n = 20) fue conformado por las aves control no inmunizadas. Todos los grupos fueron monitoreados diariamente. Los anticuerpos anti-HA de AIV en el suero fueron evaluados a los 20 días post-inmunización mediante un ELISA indirecto in house y un ensayo de inhibición de la hemaglutinación (HI, por sus siglas en inglés: hemagglutination inhibition).
- Eficacia de las vacunas frente a un desafío con AIV subtipo H5N1.
Para determinar la eficacia de las vacunas rHVT-AIV H5 e inactivada AIV (VFAR-140) contra un desafío con AIV subtipo H5N1 de alta patogenicidad, a los 21 días post-inmunización todas las aves de los grupos I, II, III, y IV fueron consideradas en el desafío. Las aves se infectaron por la vía óculo-nasal con 0,1 ml del inóculo de desafío de alta patogenicidad AIV subtipo H5N1 clado 2.3.4.b (VFAR-140) que contenía 106EID50/ave (con 0,05 ml administrados por la vía ocular y 0,05 ml por la fosa nasal). Todo el procedimiento del ensayo de desafío fue realizado dentro del laboratorio BSL-3 FARVET S.A.C. Se realizaron observaciones por un periodo de 10 días post-desafío (d.p.d) para
evaluar los signos clínicos relacionados con AIV y mortalidad, los cuales fueron registrados diariamente. Las aves muertas se retiraron inmediatamente y se auto clavaron. Asimismo, al final del experimento todas las aves sobrevivientes fueron eutanasiadas, siguiendo las indicaciones de la OIE. Finalmente, la protección fue determinada en base a la presencia de signos clínicos de AIV y mortalidad.
2.1 ELISA indirecto in-house, detección de anticuerpos anti-HA de AIV subtipo H5N1
Placas de fondo plano Nunc MaxiSorp de 96 pocillos (N° de catálogo M9410, Sigma-Aldrich) fueron recubiertas con 100 μL de proteína Influenza A H5N8 (A/duck/NY/191255-59/2002) Hemagglutinin/HA Protein (His Tag) (0.25 μg/mL) (N° de catálogo 11717-V08H, SinoBiological) diluida en tampón carbonato bicarbonato (pH: 9,6) e incubadas a 4°C durante toda la noche. Luego los pocillos fueron lavados seis veces con DPBS-Tween-20 al 0,05 % (v/v) (PBS-T 0,05 %) y se bloquearon con leche descremada DIFCO al 3 % (p/v) (N° de catálogo 232100, BD Biosciences, EE. UU.) en PBS-T al 0,05 % durante 2 h en agitación a temperatura ambiente. A continuación, las placas se lavaron seis veces con DPBS-T al 0,05 %. Se añadieron a la placa 100μL de cada muestra de suero de aves inmunizadas recolectada y diluida 1:50 con leche descremada DIFCO al 3 % (p/v) durante 1 h a 37°C. Los pocillos se lavaron seis veces con PBS-T al 0,05 % y luego se incubaron con 100μL de anticuerpo secundario Anti Chicken IgG (h&L) conjugado a HRP (N° de catálogo 60320, Alpha Diagnostic InH. Inc) diluido 1/5000 en Leche descremada DIFCO al 3 % en PBS-T al 0,05 % durante 1 h a 37 °C. Luego las placas se lavaron seis veces con PBS-T al 0,05% por última vez y se incubaron con 100 μL de 3,3′,5,5′-tetrametilbencidina (TMB) durante 15 min a temperatura ambiente. Finalmente, se detuvo la reacción agregando 50μL de H2SO4 2 N en cada pocillo y se leyeron las placas a 450 nm con un lector de microplacas EPOCH 2 (Biotek, EE. UU.). El valor del punto de corte fue calculado considerando la media de la densidad óptica de los sueros negativos más 3 desviaciones estándar.
2.2 Ensayo de inhibición de la hemaglutinación (HI)
En una placa de 96 pocillos de fondo U se depositaron 50μL de tampón DPBS más 50μL de suero en el primer pocillo y a partir de este se realizaron diluciones seriadas de 1/2, luego se añadió 50μL de AIV (VFAR-140) inactivado con formalina al 0.1 % y ajustado a un título de 4 unidades hemaglutinantes (UHA) el cual se dejó incubando durante 30 min a temperatura ambiente, luego se depositó 50μL de eritrocitos de pollo al 1% (v/v) en cada pocillo mezclando cuidadosamente, se dejó incubando durante 40 min a temperatura ambiente. Los títulos de HI fueron reportados como valores de log2.
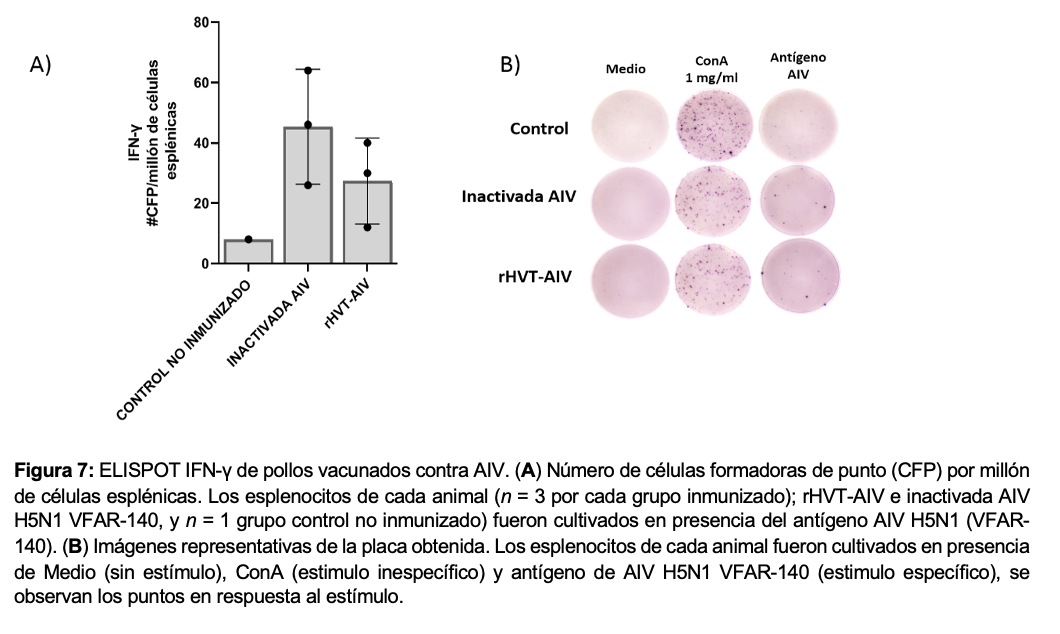
2.9. Evaluación de IFN-γ respuesta inmune celular por ELISPOT
Para determinar si las aves inmunizadas con las vacunas; rHVT-AIV H5 y la inactivada AIV H5N1 VFAR-140, adquirieron una respuesta especifica de inmunidad celular se evaluó por la técnica de CITOMETRIA DE FLUJO y ELISPOT para IFN- γ de pollo. Brevemente, en una placa de cultivo de 96 pocillos recubiertos con una membrana de PVDF, se fijaron anticuerpos de captura anti-IFN-γ de pollo, luego, se añadieron células mononucleares del bazo de las aves vacunadas, las cuales fueron estimuladas con el antígeno inactivado AIV H5N1 (VFAR-140) y finalmente fueron cultivadas a 37°C y 5% de CO2.
Posteriormente, las células fueron eliminadas mediante lavados y se colocó un anticuerpo de detección anti-IFN-γ de pollo biotinilado, el cual formó un complejo con la estreptavidina conjugada a fosfatasa alcalina, que finalmente reacciona ante el sustrato NBT/BCIP, formando un punto o spot visible en la membrana de PVDF. Los resultados se interpretaron y reportaron como cada punto o spot que corresponde a una célula secretora de IFN-γ.
3. RESULTADOS
3.1. Respuesta inmune humoral de aves inmunizadas por ELISA indirecto in-house y test de inhibición de la hemaglutinación (HI)
A los 20 días de post-inmunización, mediante el uso de un ELISA indirecto in-house, detectamos un incremento de la presencia de anticuerpos específicos contra la proteína HA de AIV H5 en los sueros de los grupos inmunizados (Grupo I con la vacuna rHVT-AIV H5, Grupo II con la vacuna AIV inactivada (VFAR-140) y Grupo III con las vacunas rHVT-AIV H5 / AIV inactivada (VFAR-140). Lo cual demostró la seroconversión. Por otro lado, en el grupo IV control no inmunizado no detectamos la presencia de anticuerpos anti-HA (Figura 6).
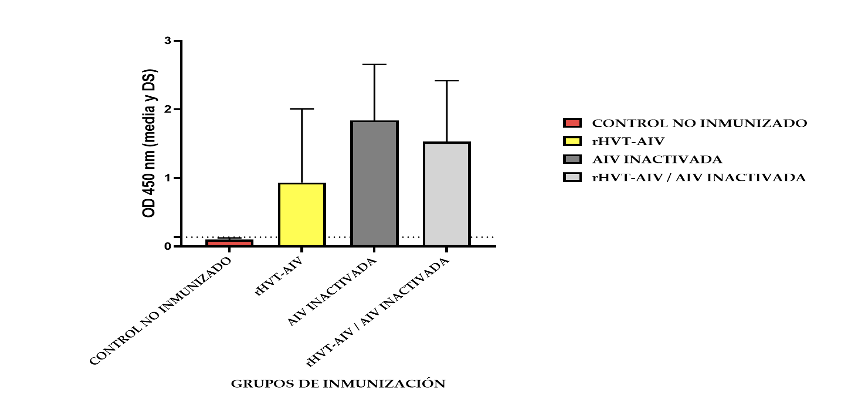
La respuesta de anticuerpos tambiénfue evaluada por un ensayo de HI contra un antígeno obtenido a partir de una cepa de AIV H5N1 aislado. A los 20 días post-inmunización el 100% de las aves vacunadas fueron positivas: el Grupo I, inmunizado con la vacuna rHVT-AIV, presentó un título de HI promedio de 1/32; el grupo II, inmunizado con la vacuna AIV inactivada, presentó un título de 1/64; el Grupo III, inmunizado con la vacuna rHVT-AIV / AIV inactivada, presentó un título de 1/64. Por otro lado, el Grupo IV, control de aves no inmunizadas, fueron negativas. (Tabla 3)

3.2. Resultados de respuesta inmune celular
Se evaluó mediante técnicas de citometría de flujo y elispot, para medir el grado de proliferación de los linfocitos de las aves vacunadas y producción de interferón gamma (IFN-y). Las pruebas demostraron una alta proliferación linfocítica y se obtuvo una buena respuesta de IFN-y en los dos grupos de aves inmunizadas con rHVT-AIV H5 y la inactivada AIV H5N1 VFAR-140 en comparación con el grupo control no inmunizado que fueron negativos. Es decir que los linfocitos de las aves vacunadas cultivados al enfrentarse con el antígeno AIV H5N1 fueron capaces de proliferarse y secretar citocinas de IFN-γ en los pocillos de cultivo, esto determina una rápida respuesta antiviral intracelular y se observa una activación de la inmunidad celular específica cuando reciben la estimulación con el antigeno AIV H5N1 (Figura 7)
3.3. Eficacia contra el desafío de AIV subtipo H5N1 de alta patogenicidad
A los 21 días post-inmunización, las aves de los grupos I, II, III, y IV fueron desafiados por vía óculo-nasal con 0,1 ml del inóculo de desafío AIV altamente patogénica subtipo H5N1 clado 2.3.4.4b (VFAR-140) con 106EID50/ave (con 0,05 ml administrados por la cavidad ocular y 0,05 ml por la cavidad nasal). Todas las aves del grupo IV control no inmunizado mostraron signos clínicos y muerte a los dos y tres d.p.d, lo que confirma la alta capacidad patogénica letal del AIV. Todas las aves inmunizadas grupos I, II, y III permanecieron clínicamente saludables durante el periodo de observación es decir libres de signos clínicos de la enfermedad AIV H5N1 (hasta 10 d.p.d), mostrando una completa protección del 100% de las aves (20/20) contra el AIV H5N1 altamente patogénica (Tabla 4). Estos resultados coinciden con los títulos de anticuerpos e inmunidad celular provocados por las vacunas; rHVT-AIV H5 e inactivada AIV subtipo H5N1 (VFAR-140) cuantificados y determinados por ELISA indirecto in-house, HI, citometría de flujo y elispot (Figura 7).

- CONCLUSIONES
Los resultados indican que las dos vacunas son capaces de conferir en aves SPF 100% de protección frente a un desafío con el virus de AIV H5N1 de alta patogenicidad clado 2.3.4.4b aislado en Perú cepa VFAR-140.
Asimismo, la vacuna viva recombinante rHVT-AIV H5 que expresa la proteína HA5 de una cepa de AIV H5N8, confirió 100% de protección contra una cepa de AIV H5N1, al igual que la vacuna inactivada AIV H5N1 VFAR-140.
Se noto un incremento en el título de anticuerpos específicos anti-H5 de AIV en los grupos I, II y III.
Se observo una respuesta inmune celular específica contra AIV H5N1 en las aves inmunizadas con ambas vacunas rHVT-AIV y la inactivada AIV H5N1 VFAR-140, esto determina y sugiere que ambas vacunas son capaces de provocar una rápida respuesta antiviral y por consiguiente existe la activación de la inmunidad humoral y celular de IFN-γ específica en caso de una infección con AIV H5N1.
Estos resultados de protección indican que la vacuna recombinante y la vacuna inactivada AIV H5N1 VFAR-140, podrían ser empleadas con éxito para el control de influenza aviar en aves de crianza comercial. (postura, carne, reproductoras etc.)
- REFERENCIAS
1. Swayne DE. 2009. Avian Influenza. Chapter 234 Avian Influenza-OIE 1–605.
2. Bruin ACM De, Funk M, Spronken MI, Gultyaev AP, Fouchier RAM, Richard M. 2022. Pathogenic Avian Influenza Virus Genesis.
3. Williams RAJ, Segovia-Hinostroza K, Ghersi BM, Gonzaga V, Townsend Peterson A, Montgomery JM. 2012. Avian influenza infections in nonmigrant land birds in Andean Peru. Journal of Wildlife Diseases 48:910–917.
4. Ghersi BM, Blazes DL, Icochea E, Gonzalez RI, Kochel T, Tinoco Y, Sovero MM, Lindstrom S, Shu B, Klimov A, Gonzalez AE, Montgomery JM. 2009. Avian influenza in wild birds, central coast of Peru. Emerging Infectious Diseases 15:935–938.
5. Nelson MI, Pollett S, Ghersi B, Silva M, Simons MP, Icochea E, Gonzalez AE, Segovia K, Kasper MR, Montgomery JM, Bausch DG. 2016. The genetic diversity of influenza A viruses in wild birds in Peru. PLoS ONE 11:1–10.
6. Ghersi BM, Sovero MM, Icochea E, Gonzalez RI, Blazes DL, Gonzalez AE, Montgomery JM. 2011. Isolation of low-pathogenic H7N3 avian influenza from wild birds in Peru. Journal of Wildlife Diseases 47:792–795.
7. Karen SH, Eliana ID, Rosa G V., Bruno G, Armando GZ. 2013. Presencia del virus de influenza aviar en aves silvestres de los humedales de Puerto Viejo, Lima. Revista de Investigaciones Veterinarias del Peru 24:98–103.
8. Kateri Bertran, Dong-Hun Lee2, Miria F. Criado, Charles L. Balzli1, Lindsay F. Killmaster, Darrell R. Kapczynski, David E. Swayne. Maternal antibody inhibition of recombinant Newcastle disease virus vectored vaccine in a primary or booster avian influenza vaccination program of broiler chickens Vaccine 36 (2018) 6361–6372
9. McMillan CLD, Young PR, Watterson D, Chappell KJ. 2021. The next generation of influenza vaccines: Towards a universal solution. Vaccines 9:1–20.
10. Palya V, Tatár-Kis T, Mató T, Felföldi B, Kovács E, Gardin Y. 2014. Onset and long-term duration of immunity provided by a single vaccination with a turkey herpesvirus vector ND vaccine in commercial layers. Veterinary Immunology and Immunopathology 158:105–115.
11. Tang N, Zhang Y, Pedrera M, Chang P, Baigent S, Moffat K, Shen Z, Nair V, Yao Y. 2018. A simple and rapid approach to develop recombinant avian herpesvirus vectored vaccines using CRISPR/Cas9 system. Vaccine 36:716–722.
12. Calderón K, Rojas-Neyra A, Carbajal-Lévano B, Luján-Valenzuela L, Ticona J, Isasi-Rivas G, Montalvan A, Criollo-Orozco M, Huaccachi-Gonzáles E, Tataje-Lavanda L, Alvarez KLF, Fernández-Sánchez M, Fernández-Díaz M, Tang N, Yao Y, Nair V. 2022. A Recombinant Turkey Herpesvirus Expressing the F Protein of Newcastle Disease Virus Genotype XII Generated by NHEJ-CRISPR/Cas9 and Cre-LoxP Systems Confers Protection against Genotype XII Challenge in Chickens. Viruses 14:793.
13. Chang P, Ameen F, Sealy JE, Sadeyen J-R, Bhat S, Li Y, Iqbal M. 2019. Application of HDR-CRISPR/Cas9 and Erythrocyte Binding for Rapid Generation of Recombinant Turkey Herpesvirus-Vectored Avian Influenza Virus Vaccines. Vaccines 7:192.
14. Xusheng Zai, Bin Shi, Hongxia Shao, Kun Qian, Jianqiang Ye, Yongxiu Yao, Venugopal Nair and AQ. 2022. Recombinant Turkey Herpesvirus Expressing H9N2 HA Gene at. Viruses 2022, Vol 14, 2495 14.
15. Balzli CL, Bertran K, Lee DH, Killmaster L, Pritchard N, Linz P, Mebatsion T, Swayne DE. 2018. The efficacy of recombinant turkey herpesvirus vaccines targeting the H5 of highly pathogenic avian influenza virus from the 2014–2015 North American outbreak. Vaccine 36:84–90.